Module 3: Elicited Behaviors and More
Module Overview
In the modules that follow this one, we will discuss associative and observational learning. Before we do that, we need to address the fact that some types of behavior are innate. To that end, we will discuss elicited behaviors. These include reflexes and modal action patterns. We will then move to a discussion of two types of non-associative learning — habituation and sensitization — and provide evidence for their adaptive advantage.
Module Outline
- 3.1. Elicited Behaviors Defined
- 3.2. Reflexes
- 3.3. Modal Action Patterns
- 3.4. Non-associative Learning
Module Learning Outcomes
- Explain what elicited behaviors are.
- Describe reflexes as a type of elicited behavior.
- Describe modal action patterns as a type of elicited behavior.
- Differentiate the two types of non-associative learning.
3.1. Elicited Behaviors Defined
Section Learning Objectives
- Define elicited behaviors.
Before we dive into behaviors that change or are acquired due to learning, we need to first discuss those that are innate and how they change. These behaviors help an organism adapt to its environment and cope with demands. Elicited behaviors are behaviors that occur due to a specific environmental stimulus. We will discuss reflexes and modal action patterns and then move to a discussion of how repeated stimulation leads to either an increase or decrease in responding or the strength of a response.
3.2. Reflexes
Section Learning Objectives
- Define reflexes.
- Exemplify reflexes in humans.
- Explain the concept of the reflex arc.
The term reflex indicates the relationship between innate behaviors and the environmental events or stimuli that elicit them. An example would be a doctor using a hammer to strike your knee (the stimulus) which causes your leg to kick out (the response). The reflex is not the response (as is commonly thought) of kicking out but the total relationship of the hammer striking the knee causing the leg to kick out. Both the stimulus and subsequent response make up a reflex. Reflexes can include a single gland or muscle such as in the case of an eye-blink in response to a puff of air while others involve several muscles or glands such as in the Moro reflex described below.
Several reflexes are exhibited by an infant as early as birth and occur automatically (they do not have to be learned). Consider the rooting reflex which consists of turning the head and sucking (response) elicited by stroking the side of the infant’s mouth or cheek (stimulus). The point is to find the nipple at feeding time. Once a nipple is in the infant’s mouth, sucking begins automatically though the coordination of the rhythmic sucking movements and breathing can be difficult for some infants at first. With time and practice, they come to handle the process fine. Another reflex present at birth is called the Moro reflex and occurs when an infant is startled by a loud sound or falls backward. The infant’s response is to throw out her arms and legs and extend her neck, which is followed by bringing her arms together. She may cry loudly. Other reflexes include walking/stepping, tonic neck, palmar grasp, and plantar grasp. These reflexes are present at birth, or close to it, showing some variation in when they appear. They help the infant survive and disappear when no longer needed. For instance, the Moro reflex disappears around 2 months after peaking around 1 month of age, the rooting reflex disappears around 4 months, while the sucking reflex stays with us for life. An infant has an instinct to step but cannot hold up her weight at first. The reflex disappears around 2 months of age but returns as the learned behavior of walking near the end of the first year. Though newborns are dependent on their caregivers, they are not completely helpless. And our reflexes change across the life span. Consider athletes for instance. How many baseball players do you see in their 40s or 50s compared to their 20s or 30s? Very few, as there is a slowing of our motor reflexes as we move from early to middle and late adulthood.
How do reflexes occur? It’s quite simple, and to make sense of it you have to understand how communication occurs in the nervous system. Consider Figure 3.1.
Figure 3.1. Communication in the Nervous System
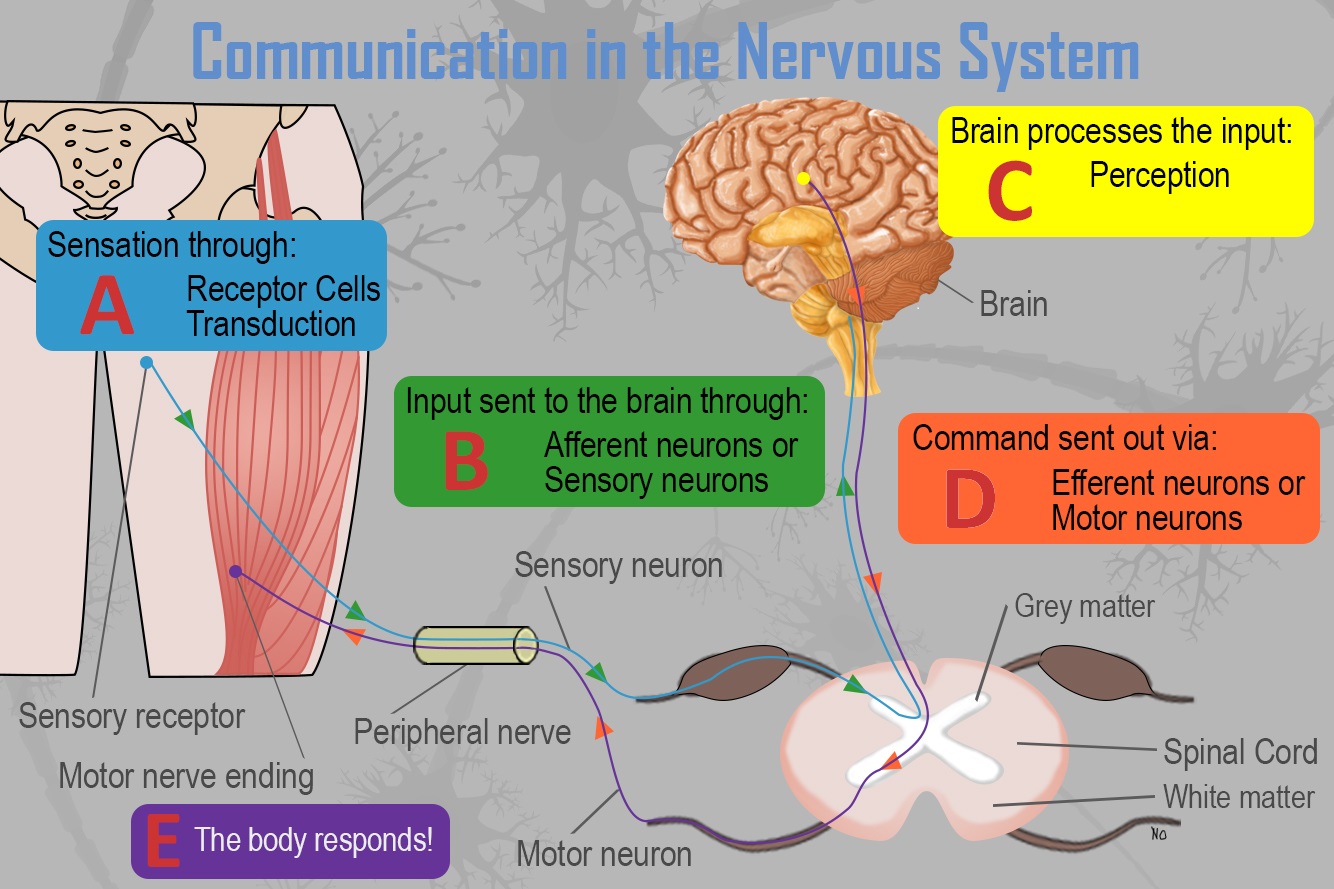
The process is best described as such:
- A. Receptor cells in each of the five sensory systems detect energy.
- B. This information is passed to the nervous system due to the process of transduction and through sensory or afferent neurons, which are part of the peripheral nervous system.
- C. The information is received by brain structures (central nervous system) and perception occurs.
- D. Once the information has been interpreted, commands are sent out, telling the body how to respond (Step E), also via the peripheral nervous system.
With reflexes, when the doctor strikes your knee with the hammer, receptor cells in your skin detect this energy and pass the information to your nervous system. Afferent or sensory neurons then carry this information to the brain. Once in the brain, it is processed and commands are sent out via efferent or motor neurons telling your knee to kick out. It may be quicker than this as the information could produce a more immediate response via the action of interneurons. How so?
Consider a withdrawal reflex that may be needed if you touch a hot plate or stove (stimulus). You will pull your hand back pretty quickly (response) as the stimulus is painful. This process occurs in three steps. First, receptor cells in the skin detect pressure, warmth, cold, light touch, or in this case, pain. This information is sent to the brain as described above but the central nervous system (CNS) can distinguish between the various pieces of sensory information it is receiving since different pathways are activated due to different stimuli. Excited afferent neurons carry the pain information toward the brain, but they are intercepted in the spinal cord by excitatory interneurons that in turn stimulate efferent neurons, resulting in the muscle in the arm flexing or bending the elbow joint. This moves our hand away from the hot stove. Second and simultaneously, the afferent neurons also activate inhibitory interneurons which prevent the efferent neurons from sending commands for the triceps to contract which would result in the arm straightening out, putting us back in harm’s way. This reflex includes the stimulation of nerves in one muscle while inhibiting the nerves in another, opposing muscle to create the desired response, called reciprocal innervation. Finally, the afferent neuron stimulates other interneurons which continue moving the original message from the spinal cord to the brain itself so that we are made aware of the pain, where it occurred, and what caused it. This will lead to a decision about what to do and we can store away the information in memory so that in the future we are more cautious when moving near a hot stove.
Taken together, the examples above show a relatively simple process called a reflex arc, which demonstrates the earlier point that a reflex is a process, not the terminal behavior. Let’s now tackle a more complex process.
3.3. Modal Action Patterns
Section Learning Objectives
- Define and describe modal action patterns.
- Exemplify modal action patterns.
A more complex type of innate response is called a modal action pattern (MAP) and can be specific to an individual species (Barlow, 1977). Be advised that in the literature the term fixed action pattern is also used, but is less common, and MAPs have been referred to as instincts in the past. They are similar to reflexes because they are relatively stable across individuals and situations, and as such are described as highly stereotyped. They also have a genetic component and are caused by specific stimuli. The difference is that they are fairly complex, involve the entire organism and not specific muscles like the neck or fingers, and can vary. MAPs are elicited by releasing stimuli, or sign stimuli, in the environment, and aid a species’ survival by dealing with threats, finding food sources, and passing genetic information from one generation to the next. Once began, the MAP must be carried out to completion.
Ethologists have uncovered many such MAPs and their sign stimulus. For instance, a nesting Graylag Goose will roll a displaced egg back to its nest using its beak. The goose will continue with this behavior even if the egg is removed during the behavior. It is as if the egg is still there. The sight of the displaced egg is the sign stimulus that brings about or elicits the behavior of egg-retrieval (rolling it back in the nest).
Niko Tinbergen (1952) described the courtship behavior of male three-spined stickleback fish during mating season, which begins by establishing a territory, building a nest in it, and changing the color of its underbelly to red. Males entering their territory are attacked while females are courted and encouraged to enter the nest and lay eggs. The presence of the other male is not the sign stimulus for aggressive behavior, but the sight of its red underbelly. Similarly, aggressive behavior is displayed by a male bighorn sheep who wins its mate by bashing its head against that of a competing male. In the case of some birds, females decide which male to mate with based on how elaborate its plumage is and its dance. The male knows to display its plumage and dance (the response) at the sight of the female (the sign stimulus).
Kelp gull chicks cause their mother to regurgitate by pecking (the response) a red spot they see on their mother’s beak (the sign stimulus). Moths are also known to instantly fold their wings and drop to the ground (the response) if they detect ultrasonic signals produced by bats (the stimulus).
Interestingly, some species have evolved to exploit the MAPs of another species by mimicking its sign stimulus. Called code-breaking, one species duplicates another species releasing mechanism. For instance, the North American cowbird, classified as a brood parasite, lays its eggs in another species nest and once hatched, its young push the eggs and hatchlings of the other species out of the nest leaving them to die. Being that it is larger and louder than the original species, it causes its “adoptive” parent to forage more frequently than it would normally need to and to feed the impostor baby.
3.4. Non-associative Learning
Section Learning Objectives
- Contrast habituation and sensitization.
- Describe habituation.
- Clarify how tolerance is a form of habituation.
- Describe sensitization.
- Clarify how the dual process theory explains habituation and sensitization.
- Clarify how the opponent process of emotion shows the malleability of our emotions.
- Explain where there is an adaptive advantage to habituation and sensitization.
As noted above, repetitive stimulation can lead to either an increase or decrease in our likelihood of responding or the intensity of our response. When the size or probability of a response decreases, habituation is occurring, such as when we no longer hear the ticking of a clock or the fan in our computer. We may also experience an increase in our responding or its intensity, called sensitization. A police officer involved in a shoot-out will not habituate to the sound of gunshots, but actually become more aware of them.
These two forms of learning are non-associative in nature, or not learned by linking together environmental events, as respondent and operant conditioning are.
3.4.1. Habituation
Habituation is not to be confused with sensory adaptation, which occurs when a sense organ becomes disabled, such as being temporarily blinded by a camera flash or having your hearing limited due to a loud sound like a gun being fired on a range. It should also not be confused with response fatigue or when a muscle is incapacitated due to repeated use such as weight training one’s legs and then not being able to run in an emergency. Habituation occurs within the nervous system while sensory adaptation and response fatigue occur outside it and in sense organs and muscles, respectively. It is a nearly universal occurrence in the animal kingdom and helps organisms save time and energy by not engaging in behaviors that are not functional.
Habituation generally ends if the stimulus is not present for a period of time and can either be short-term or long-term, indicating the length of time or durability of the habituation effect. When we think about learning, long-term habituation is what we are describing as it is a relatively permanent and stable change in behavior, as the definition of learning from Module 1 stated. The response slowly decreases with repeated stimulation and our ability to respond slowly recovers when the stimulus ends. In contrast, in short-term habituation, the response quickly decreases with repeated exposure to the stimulus and quickly recovers once gone.
Drug tolerance represents a form of habituation. According to the National Institute of Drug Abuse (NIDA), tolerance occurs when a drug is used repeatedly over time. The person no longer responds to the drug the way he or she initially did, meaning that higher doses of the drug are needed to obtain the same effect achieved early on. According to drugabuse.com, tolerance can take three forms. Acute, or short-term, tolerance results from repeated exposure to a drug over a short period of time while chronic, or long-term, tolerance develops when a person adapts to constant exposure to the drug over weeks or months. Finally, learned tolerance may result from frequent exposure to the drug. People who abuse alcohol for months or years may not appear intoxicated to others because they learn to compensate for the effects of alcohol on their coordination by practicing a task repeatedly while intoxicated. This tolerance disappears if the task is altered (Vogel-Sprott, 1997). Kesner and Cook (1983) found that 96 male Long-Evans rats developed tolerance to morphine in a nondistinctive environment due to repeated presentation of the drug, and that tolerance was quicker in massed rather than spaced presentations of the morphine.
Dishabituation occurs when an organism’s state of arousal is enhanced, leading to an increase in the response that previously was habituated. If you habituated to the sound of the clock ticking in your office, you might notice it again if an unrelated, but novel, stimulus presents itself such as a coworker talking to you. Dishabituation represents a separate process from sensitization.
3.4.2. Sensitization
As with habituation, sensitization ends when the stimulus is not present for a period of time. Unlike habituation, sensitization can generalize to similar stimuli. Consider that soldiers returning from war will jump at the sound of a car backfiring as it sounds like an explosion caused by an artillery shell or IED (Improvised Explosive Device).
Sensitization is also important in sexual behavior and responding to tactile stimuli such as kissing or hugging your significant other. When doing so, you become aroused, unlike when you engage in similar behaviors with your mother or a sibling. For instance, one study showed that vibrotactile sensitivity increased in a sample of 30 heterosexual, healthy males after viewing an erotic video, but did not occur in response to a non-erotic video (Jiao et al., 2007).
3.4.3. An Explanation for Habituation and Sensitization
The two processes of habituation and sensitization are controlled by different mechanisms that are described by the dual-process theory (Groves & Thompson, 1970). Habituation occurs due to the S-R system or through changes in neurons controlling our response. The S-R system is best represented by the reflex arc, and subsequent activation of this system causes the build-up of habituation. It is active with each occurrence of an eliciting stimulus.
On the other hand, sensitization occurs due to changes in an organism’s arousal, called the state system, which leads to excitability. The state system consists of those parts of the nervous system involved in our readiness to respond to an environmental event, and only activates due to arousing events.
3.4.4. The Malleability of Emotional States
Given what you have read so far, would it surprise you to learn that emotional states can also change due to the repeated exposure to environmental stimuli? The opponent process theory of emotion (Solomon & Corbit, 1974) states that our primary emotional reaction to an emotion-arousing stimulus, or a process, is followed by an opposite after-reaction, or b process, which counteracts this shift, and the primary reaction becomes weaker or habituates with repeated stimulation, while the after-reaction becomes stronger. This serves to maintain homeostasis or to keep our emotions fairly even or neutral. We seek to minimize highs and lows. For instance, in the first few stimulations opiate users will experience euphoria, a rush, and pleasure (a process) which will be followed by craving and aversive withdrawal signs for a short duration (b process). But after many such stimulations, they will experience a loss of euphoria, normal feelings, and no rush (a process), followed by an intense craving and agony lasting a long time, which is called abstinence agony (b process). Solomon and Corbit (1974) refer to this behavioral phenomenon of addiction as a “ghastly experience” (pg. 123).
3.4.5. Are Habituation and Sensitization Adaptive?
To understand the adaptive value of habituation and sensitization, we need to understand the role of stimulus intensity. Basically, low-intensity stimuli lead to habituation while high-intensity stimuli lead to sensitization. Why is that? Stimuli that are low intensity tend to be non-threatening, such as the fan in your computer running, while stimuli that are high intensity tend to be threatening, such as the sound of a gun firing. What is important is the threshold level of the organism when the first stimulus in a series occurs such that the greater the response the lower the organism’s sensory threshold prior to the stimulus. This initial response level will determine whether habituation or sensitization occurs (Eisenstein, Eisenstein, & Smith, 2001).
And habituation leads to a preservation of valuable energy that can be better spent by the organism. This represents the ability of an organism to change its behavior due to experience, which reflects learning.
Module Recap
Module 3 discussed forms of innate behavior to include reflexes and modal action patterns, and our inherent predisposition to respond more or less to recurring stimuli in our environment based on our threshold level when the stimulus is first encountered. We now move to associative and observational learning in Modules 4-8.
2nd edition
